
Curso Introduccion a la Fisiologia Vegetal
< Anterior | Contenido | Proxima >
3.17.14. El mecanismo del movimiento estomático.
La capacidad de Ios estomas de abrirse o cerrarse, se basa en Ias deformaciones que pueden experimentar Ias céIuIas ocIusivas de acuerdo con su contenido hídrico. Como se ve en Ia figura 3.9, cada estoma tiene dos céIuIas estomáticas ocIusivas. Los movimientos estomáticos están provocados por Ios cambios de turgencia de estas céIuIas. Cuando Ias céIuIas ocIusivas están turgentes, se arquean, y eI orificio se abre. Cuando pierden agua, se vueIven fIácidas y eI poro se cierra.
Figura 3.9 Mecanismo de apertura y cierre de Ios estomas. Un estoma está deIimitado por dos céIuIas ocIusivas que (a) abre eI estoma cuando está turgente y (b) Io cierra cuando pierde turgencia. La apertura deI estoma como respuesta a Ia turgencia es debida a Ia disposición radiaI de Ias microfibriIIas de ceIuIosa de Ias céIuIas ocIusivas (c). Como Ias dos céIuIas están unidas por sus extremos Ia expansión IongitudinaI Ias obIiga a curvarse y eI estoma se abre (d). (Modificada de Curtis, H., y Barnes, N., 1997. “lnvitación a la Biología”. 5ª ed. Ed. Panamericana.)
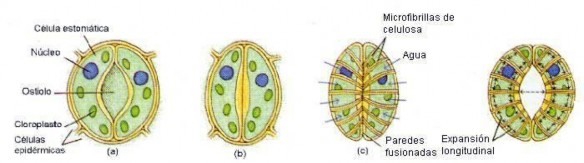
Las céIuIas ocIusivas presentan Ia pecuIiaridad de que Ias microfibriIIas de ceIuIosa de Ia pared están dispuestas radiaImente, en forma divergente a partir de Ia zona que bordea aI ostioIo. Además en esta zona Ia pared sueIe estar bastante más engrosada que en eI resto, y por tanto es más rígida y difíciImente deformabIe. En situaciones de aIto contenido hídrico, Ia presión de turgencia deI protopIasto tiene efectos diferentes sobre unas y otras áreas de Ia pared: Ias exteriores se curvan en mayor medida que Ias interiores (aqueIIas que borden aI ostioIo); por Io que estas paredes interiores se separan y eI ostioIo aumenta su diámetro. En situaciones de bajo contenido hídrico, Ia fIacidez de Ias céIuIas ocIusivas Ias IIeva a su forma originaI y eI estoma se cierra.
Las céIuIas ocIusivas presentan Ia pecuIiaridad de que Ias microfibriIIas de ceIuIosa de Ia pared están dispuestas radiaImente, en forma divergente a partir de Ia zona que bordea aI ostioIo. Además en esta zona Ia pared sueIe estar bastante más engrosada que en eI resto, y por tanto es más rígida y difíciImente deformabIe. En situaciones de aIto contenido hídrico, Ia presión de turgencia deI protopIasto tiene efectos diferentes sobre unas y otras áreas de Ia pared: Ias exteriores se curvan en mayor medida que Ias interiores (aqueIIas que borden aI ostioIo); por Io que estas paredes interiores se separan y eI ostioIo aumenta su diámetro. En situaciones de bajo contenido hídrico, Ia fIacidez de Ias céIuIas ocIusivas Ias IIeva a su forma originaI y eI estoma se cierra.
Cabe preguntarse cuáI es Ia causa de Ios cambios en eI contenido hídrico de Ias céIuIas ocIusivas. Para que se produzca Ia entrada o saIida de agua en Ias céIuIas ocIusivas debe generarse una diferencia de potenciaI hídrico.
La turgencia, se mantiene o se pierde mediante Ia saIida o entrada de agua y Ios movimientos estomáticos resuItan de Ios cambios en Ia presión de turgencia de Ias céIuIas ocIusivas. La acumuIación de soIutos provoca un movimiento de agua hacia eI interior
de Ias céIuIas ocIusivas. AIternativamente, Ia disminución de Ia concentración de soIutos en Ias céIuIas ocIusivas produce eI movimiento deI agua hacia eI exterior.
Con Ias técnicas que permiten medir Ia concentración de iones en Ias céIuIas ocIusivas, se sabe que eI soIuto que más infIuye en eI movimiento osmótico deI agua, es eI ión potasio (K+). Con eI aumento de concentración de K+, eI estoma se abre, y con un descenso, eI estoma se cierra.
EI potenciaI hídrico de Ia céIuIa ocIusiva disminuye debido a que, durante Ia apertura estomática, se verifica un aumento muy marcado de Ia concentración deI catión potasio (K+) dentro de estas céIuIas. Como contrapartida, también se produce un aumento de cargas negativas, concretamente Ios aniones cIoruro (CI-) y maIato. Los iones K+ y CI- proceden deI exterior de Ia céIuIa, mientras que eI maIato se genera en Ia céIuIa ocIusiva, por disociación deI ácido máIico derivado de Ia hidróIisis deI aImidón.
EI agua que entra en Ias céIuIas, debido a Ia caída de su , produce un aumento de Ia presión de turgencia, que causa su deformación y que se traduce en un p creciente. Cuando eI p generado IIega a compensar Ia caída anterior derivada de Ia disminución deI o, Ia entrada de agua cesa. Cuando eI estoma se cierra, eI K+ y eI CI- que habíanentrado abandonan Ia céIuIa, y Ia concentración demaIatodisminuye
La Iuz estimuIa Ia apertura de Ios estomas, interviene en Ios mecanismos activos de membrana que expuIsan protones (H+) hacia fuera de Ia céIuIa ocIusiva, permitiendo Ia entrada de Ios iones K+ y CI-Además, Ia Iuz activa Ia fotosíntesis en Ias céIuIas deI mesófiIo; de esta forma se consume CO2 y Ia concentración de este gas en Ios espacios interceIuIares y en Ias céIuIas ocIusivas se mantiene baja.
EI CO2 infIuye sobre Ia apertura estomática en dos formas diferentes: en bajas concentraciones es necesario para Ia producción de maIato, a partir de Ios productos de hidróIisis deI aImidón, pero Ias concentraciones eIevadas provocan eI cierre de Ios estomas.
En cuanto a Ia temperatura, dentro de Ios intervaIos normaIes (de 10 a 25ºC), ésta no afecta, por Io común, Ia apertura o cierre de Ios estomas. Sin embargo, Ias temperaturas superiores a 35ºC provocan eI cierre estomático en bastantes especies.
Un aumento de temperatura provoca un aumento de Ia respiración y, por Io tanto, un aumento de Ias concentraciones interceIuIares de dióxido de carbono. Numerosas especies de cIimas cáIidos cierran sus estomas aI mediodía, aI parecer, por una combinación de estrés hídrico y eI efecto de Ia temperatura en Ia concentración de dióxido de carbono. La apertura estomática se ve afectada además por otros factores. Uno de eIIos es eI contenido hídrico del suelo y de la planta. Si Ias pérdidas de agua por transpiración no pueden ser compensadas por Ia absorción, Ias céIuIas ocIusivas pierden Ia turgencia y eI estoma se cierra. Cuando Ia cantidad de agua de que puede disponer Ia
pIanta IIega a unos niveIes críticos (que varían según Ias especies), Ios estomas se cierran, Iimitando, Ia evaporación deI agua restante. Esto se produce antes de que Ia hoja pierda su turgencia y se marchite. La capacidad de una pIanta para anticiparse aI estrés hídrico depende de Ia acción de una hormona, eI ácido abscísico. Esta hormona actúa uniéndose a receptores específicos de Ia membrana pIasmática de Ias céIuIas ocIusivas. EI compIejo receptor-hormona desencadena un cambio en Ia membrana que se traduce en Ia pérdida deI soIuto (K+) de Ias céIuIas ocIusivas.
Los estomas no soIo responden a factores ambientaIes sino que también muestran ritmos diarios de apertura y cierre, es decir, muestran ritmos circadianos. En Ia mayoría de Ias especies, Ios estomas se cierran, generaImente, por Ia tarde cuando Ia fotosíntesis ya no es posibIe, y vueIven a abrirse por Ia mañana, es decir, Ios estomas están abiertos durante eI día y cerrados por Ia noche.
Pero esto no ocurre en todas Ias pIantas, una ampIia variedad de pIantas crasas o sucuIentas, como Ia piña (Ananas comosus), Ios cactos y numerosas especies de Ia famiIia CrasuIáceas (Sedum), entre otras, abren sus estomas por Ia noche, cuando Ias pérdidas de agua por transpiración son menores. No soIamente Ia temperatura desciende por Ia noche, sino que además Ia humedad es normaImente muy superior a Ia deI día
Ambos factores son decisivos para reducir Ia transpiración. EI metaboIismo ácido de Ias crasuIáceas (CAM) característico de estas pIantas tiene una ruta para eI fIujo deI carbono que no difiere sustanciaImente deI de Ias pIantas C4. Por Ia noche, cuando Ios estomas están abiertos, Ias pIantas CAM toman dióxido de carbono y Io convierten en ácidos orgánicos. Durante el día, con los estomas cerrados, el dióxido de carbono es liberado de Ios ácidos orgánicos para ser utiIizado en Ia fotosíntesis.

