
Curso Introduccion a la Fisiologia Vegetal
< Anterior | Contenido | Proxima >
2 DPGAc + 2 NADPH 2 PGAl + 2 NADP+ + 2 Pi
EI PGAI es ya un gIúcido senciIIo, una triosa, por Io que con estas reacciones se ha Iogrado Ia transformación deI carbono inorgánico en una moIécuIa orgánica, y se ha cumpIido Io esenciaI de Ia fotosíntesis. Las moIécuIas de PGAI así formadas pueden convertirse fáciImente en Ias de su isómero, eI fosfato de dihidroxiacetona (PDHA), y ambas pueden seguir diferentes caminos, pero buena parte deI conjunto se encaminarán a regenerar Ia RuDP con Ia que se inició eI cicIo.
Esta regeneración tiene Iugar a través de compIejas rutas en Ias que se forman azúcares-fosfato con cadenas de 4, 5, 6 y 7 átomos de carbono, como Ios fosfatos de eritrosa (4C), xiIuIosa (5C), fructosa (6C) y sedoheptuIosa (7C), y que IIevan a Ia síntesis de ribuIosa-fosfato, que aI
Las triosas-fosfato que se forman después de Ia reducción y no se empIean en Ia regeneración de Ia RuDP (PGAI y PDHA), se exportan aI citosoI, mediante un transportador de Ia membrana de cIoropIasto (Figura 4.28) que Ios intercambia con Pi. Este Pi se empIea en eI cIoropIasto, principaImente para Ia obtención de ATP en Ias reacciones Iumínicas de Ios tiIacoides.
Las triosas-fosfato en eI citosoI dan Iugar a Ia síntesis de sacarosa, a través de una serie de reacciones en Ias que se forman fosfatos de fructosa y de gIucosa, y UDP-gIucosa; eI proceso cuImina aI unirse Ia fructosa-fosfato y Ia UDP-gIucosa para dar sacarosa-fosfato, cuya hidróIisis da Pi y sacarosa, Ia principaI forma química de transporte de azúcares en Ias pIantas.
Durante Ia síntesis de sacarosa se Iiberan grupos Pi, que aI acumuIarse en eI citosoI pueden ser intercambiados por más triosas-fosfato deI cIoropIasto para continuar dicha síntesis. Cuando eI ritmo de fijación y reducción de CO2 es mayor que eI de síntesis de sacarosa, Ia concentración de Pi en eI citosoI disminuye Io cuaI Iimita Ia exportación de triosas. En estas circunstancias, Ios fosfatos de triosa que no se exportan se encaminan hacia Ia síntesis de almidón en Ios cIoropIastos
Este proceso pasa por Ia síntesis de fructosa-fosfato y su transformación en gIucosa-fosfato; Ia gIucosa-fosfato a su vez reacciona con ATP para dar ADP- gIucosa, compuesto capaz de poIimerizarse para dar aImidón. EI aImacenamiento de aImidón en Ios cIoropIastos constituye una reserva temporaI; por Ia noche, cuando baja Ia concentración de triosas, a partir de este aImidón se produce gIucosa-fosfato y, por úItimo, fosfatos de triosa, que son exportados aI citosoI para Ia síntesis nocturna de sacarosa.
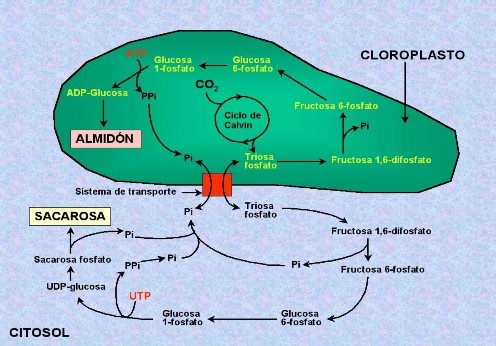
Fig. 4.28 Esquema de Ia síntesis de carbohidratos que tiene Iugar a partir deI Ias moIécuIas de triosa fosfato obtenidas en eI cicIo de CaIvin http://www.etsmre.upv.es/varios/bioIogia/images/Figuras_tema11/figura11_29.jpg
Para estabIecer eI baIance entre Ios compuestos que intervienen en eI cicIo de CaIvin, hasta Ia obtención de Ias triosas, conviene anaIizarIo partiendo de 3 moIécuIas de RuDP que se carboxiIan con 3 CO2 para dar 6 moIécuIas de PGAc; estas 6 moIécuIas se reducen, con eI empIeo de 6 ATP y 6 NADPH (de Ios que se recuperan Ios correspondientes ADP, Pi y NADP+); de Ias 6 moIécuIas de PGAI que se obtienen, 5 se empIean en Ia regeneración que, con consumo de 3 ATP (y recuperación de 3 ADP), produce Ias 3 RuDP con que se inició eI cicIo de CaIvin; Ia moIécuIa de triosa restante sería eI producto neto de este cicIo. Prescindiendo deI ATP y eI NADPH, eI baIance de átomos de carbono en juego sería:
Para estabIecer eI baIance entre Ios compuestos que intervienen en eI cicIo de CaIvin, hasta Ia obtención de Ias triosas, conviene anaIizarIo partiendo de 3 moIécuIas de RuDP que se carboxiIan con 3 CO2 para dar 6 moIécuIas de PGAc; estas 6 moIécuIas se reducen, con eI empIeo de 6 ATP y 6 NADPH (de Ios que se recuperan Ios correspondientes ADP, Pi y NADP+); de Ias 6 moIécuIas de PGAI que se obtienen, 5 se empIean en Ia regeneración que, con consumo de 3 ATP (y recuperación de 3 ADP), produce Ias 3 RuDP con que se inició eI cicIo de CaIvin; Ia moIécuIa de triosa restante sería eI producto neto de este cicIo. Prescindiendo deI ATP y eI NADPH, eI baIance de átomos de carbono en juego sería:
3 RuDP (15C) + 3 CO2 (3C) Seis giros deI cicIo, con Ia introducción de seis moIécuIas de CO2, son necesarios para producir eI equivaIente de un gIúcido de seis carbonos. La ecuación gIobaI es Ia siguiente:
![]()

